1. Axon Guidance
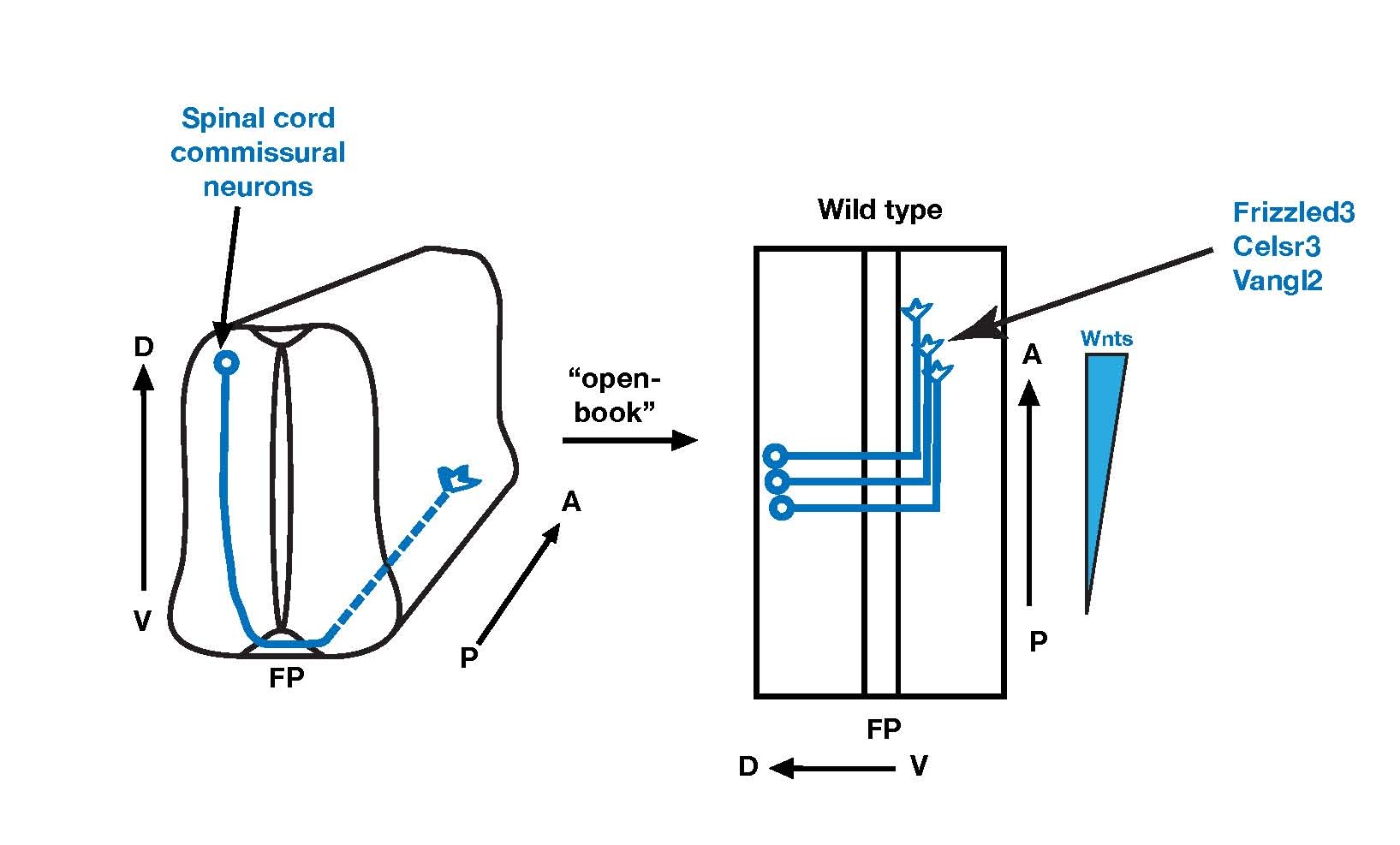
We pioneered the finding that Wnt family proteins provide key directional information along the major axes of the nervous system for axon pathfinding and topographic projections. We are currently studying how these guidance cues are laid out in concentration gradients and how growth cones recognize and respond to these gradients. We were the first to show that the conserved apical-basal and planar cell polarity pathways mediate Wnt signaling in growth cone turning. We found that apical-basal polarity signaling component, aPKC, promotes the endocytosis of a planar cell polarity component, Frizzled3, and are testing whether it is part of an amplification mechanism for asymmetric signaling to polarize growth cones.
2. Circuit repair after traumatic injury
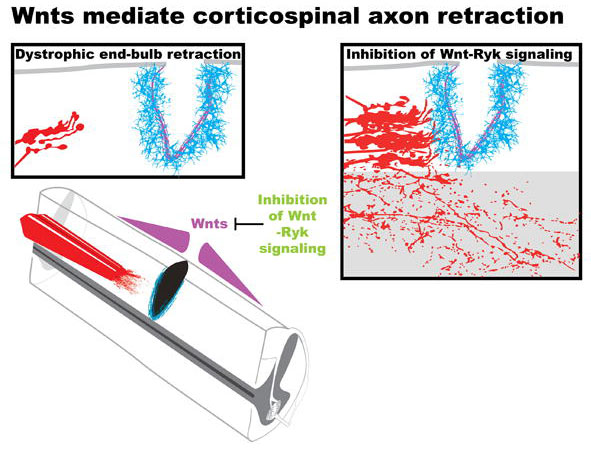
Traumatic injuries of the brain and the spinal cord lead to loss of many important functions. We found that the Wnt signaling pathways are reactivated after spinal cord injury. The reinduced Wnt inhibitory system limits motor and sensory axon growth and functional recovery. Our recent studies showed that combining molecular manipulation to enhance axon plasticity with behaviorally-guided training offers the maximal functional recovery. We are currently studying the network basis of neural circuit remodeling after spinal cord injury and traumatic brain injury.
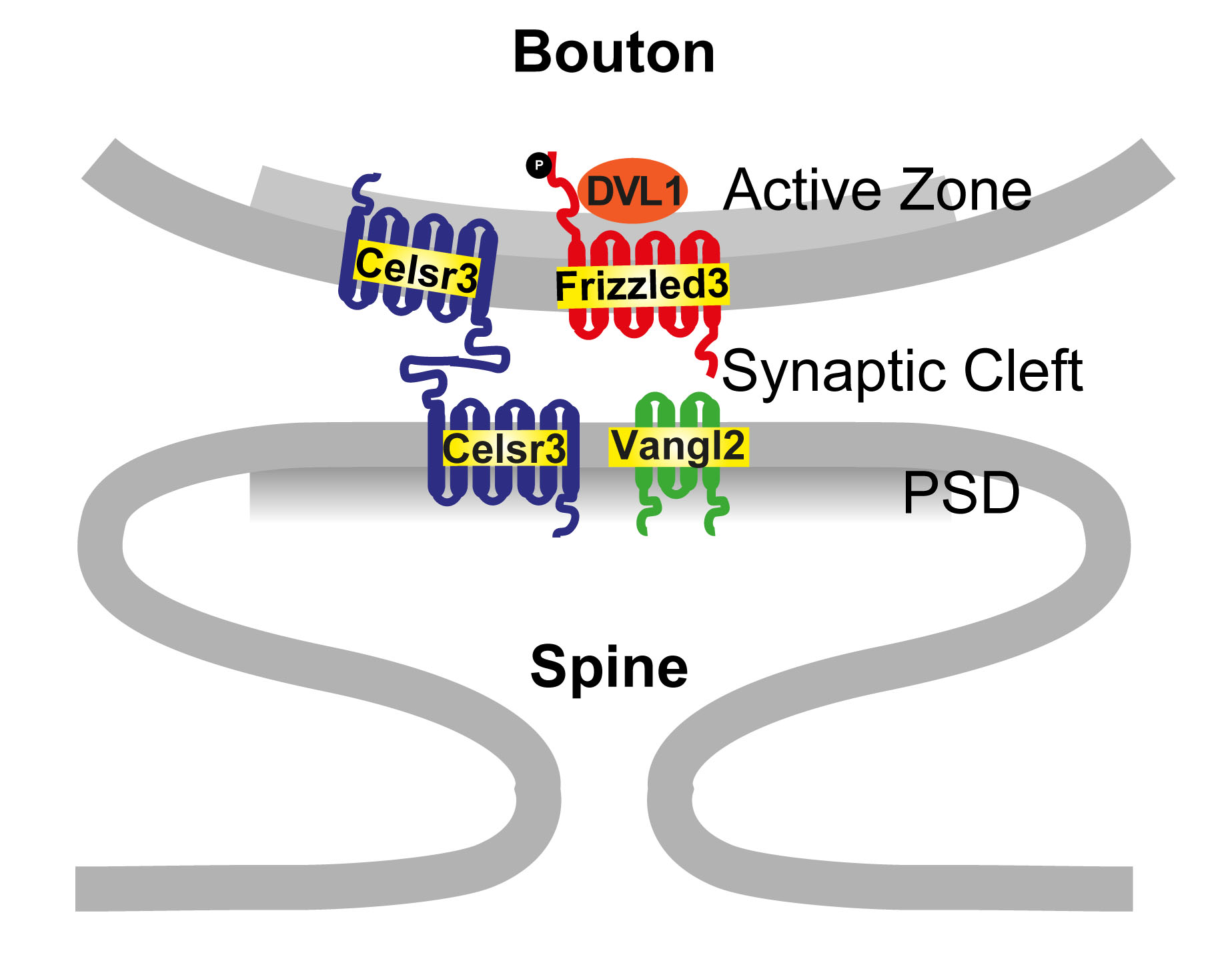
3. Synapse formation and neurodevelopmental disorders
Our recent finding suggests that planar cell polarity signaling components are present in developing glutamatergic synapses and play essential roles in glutamatergic synapse formation. This opens up new opportunities to study synaptogenesis and plasticity, fundamental to understanding neural circuit function. Mutations in planar cell polarity genes, such as the Prickles, are associated with neurodevelopmental disorders, such as autism and epilepsy. We are studying the molecular and circuit mechanisms of these diseases and exploring strategies for therapeutic interventions.