![]()
| Flower Development |
|
 Arabidopsis
flowers originate as small groups of undifferentiated cells called flower meristems on the flanks of the inflorescence (shoot) meristem. Arabidopsis
flowers originate as small groups of undifferentiated cells called flower meristems on the flanks of the inflorescence (shoot) meristem.In contrast to the indeterminate shoot meristem which essentially gives rise to flower primordia indefinitely, flowers are determinate structures that produce a defined number of organ primordia. |
|
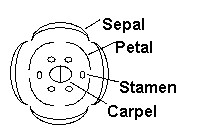
| These organ primordia reside
in four concentric rings called whorls, with four sepals in the outermost first whorl,
followed by four petals, six stamens, and two fused carpels in the 2nd, 3rd,
and 4th whorls, respectively. There are numerous genes required for the initiation and development of flowers, and for simplicity they can be divided into distinct classes. |
|
| The first class are Flowering Time genes, mutations in which cause early or
late flowering. Flowering Time genes can themselves be divided into distinct classes,
based on their differential responses to a number of environmental conditions, such as day
length and vernalization. (See review by Weigel, Ann. Rev. Genet. 29:19-39, 1995
and references therein.) The second class specifies Meristem Identity, and includes genes such as LEAFY, APETALA1, and CAULIFLOWER which specify flower meristem identity, as well as TERMINAL FLOWER which maintains inflorescence meristem identity. |
Wildtype flower |
| A third class
includes the Flower Organ Identity genes,
which determine the fate of organ primordia and are incorporated into the "ABC" model
of flower development. Examples of organ identity genes include APETALA1 (which
is involved in both meristem and organ identity), APETALA2,
APETALA 3, PISTILLATA and AGAMOUS. A fourth class includes late-acting genes that control ovule development and extensive genetic and recent molecular studies have begun to uncover the complex array of interactions among genes in this class. |
Related Links
|
![]()