| Flower organ identity |
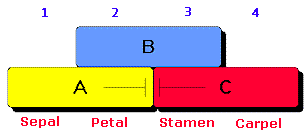
| A great deal of attention has focused on the genes that specify organ identity, leading to the "ABC" model of flower development. This model proposes that at around the time of organ initiation, the flower meristem is partitioned into three overlapping fields of gene activity, with each field defining two adjacent whorls. | Related Links |
Click on any of the three activities shown above to see more information about the appropriate class of floral organ identity genes
|
|
| Field A encompasses whorls 1
and 2; field B encompasses whorls 2 and 3; Field C encompasses whorls 3 and 4. The model
suggests that the A and C functions specify sepals and carpels, respectively, whereas the
combined activities of AB and BC specify petals and stamens, respectively. The model further suggests that the A and C functions are mutally antagonistic, such that in a-function mutants, the C domain expands to include all whorls, and similary in c-function mutants, the A domain expands to include all whorls. The A function requires at least two genes, APETALA1(AP1) and APETALA2(AP2), the B function requires at least two genes, APETALA3(AP3) and PISTILLATA(PI), and the C function requires AGAMOUS(AG). As predicted by the model, the A function gene AP1, which is normally expressed in whorls 1 and 2, is expressed on all whorls of ag mutants. Similarly, the AG gene is normally expressed only in whorls 3 and 4, but is expressed in whorls 1 and 2 in ap2 mutants. |
|
| Although this model is simplistic in nature, it is
generally applicable to a wide variety of distantly related plant species. This
generalization is based on the fact that similar genes, expression patterns and mutant
phenotypes occur in diverse plants, and from the observation that genes from one species
produce the predicted phenotypes when ectopically expressed in a distantly related
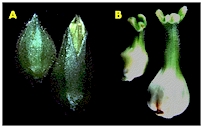
species. Shown here is a photo of transgenic tobacco overexpressing the Brassica AG gene. |
A. wildtype tobacco flowers B. 35S BAG tobacco flowers have a sepal to carpel conversion |
| Selected References:
Bowman, J.L., Smyth, D.R., and Meyerowitz, E.M. (1991). Genetic interactions among floral homeotic genes of Arabidopsis. Development 112, 1-20. Coen, E.S., and Meyerowitz, E.M. (1991). The war of the whorls: genetic interactions controlling flower development. Nature 353, 31-37. Coen, E.S., Romero, J.M., Doyle, S., Elliot, R., Murphy, G., and Carpenter, R. (1990). FLORICAULA : homeotic gene required for flower development in Antirrhinum majus. Cell 63, 1311-1322. Haughn, G.W. and Somerville, C.R. (1988) Genetic control of morphogenesis in Arabidopsis. Dev. Genet. 9:73-89. Mandel, A.M., Bowman, J.L., Kempin, S.A., Ma, H., Meyerowitz, E.M., and Yanofsky, M.F. (1992b). Manipulation of flower structure in transgenic tobacco. Cell 71:133-143. Schwarz-Sommer, Z., Huijser, P., Nacken, W., Saedler, H., and Sommer, H. (1990). Genetic control of flower development: homeotic genes in Antirrhinum majus. Science 250, 931-936. Weigel, D. and Meyerowitz, E.M. (1994) The ABCs of floral homeotic genes. Cell 78:203-209. Weigel, D. (1995) The genetics of flower development: From floral induction to ovule morphogenesis. Ann. Rev. Genet. 29:19-39. Yanofsky, M.F. (1995) Floral meristems to floral organs: Genes controlling early events in Arabidopsis flower development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 46:167-188. |