
Welcome to Kaustuv Roy’s research group at the University of California San Diego. Our research is focused on better understanding: (i) the processes that generate and maintain large scale spatial gradients in biodiversity (ii) how such gradients are being impacted by climate change and other anthropogenic impacts (e.g. human harvesting) and (iii) the ecological and evolutionary consequences of such changes. The research in the lab is interdisciplinary and we address these questions using data for coastal marine invertebrates as well as the microbial taxa associated with them (i.e. their microbiomes).
More about the projects we are working on …….
Origin and maintenance of the Latitudinal Diversity gradient
 The latitudinal diversity gradient (LDG) – a dramatic increase in the richness of species and higher taxa from the poles to the tropics – is a global biodiversity pattern shared by both marine and terrestrial ecosystems. Understanding the processes responsible for the origin and maintenance of the LDG has been a focus of biology for over two centuries and is essential for formulating effective management and conservation strategies. Yet, despite considerable progress, the environmental and biological drivers of the LDG still remain poorly understood; two dozen major hypotheses have been proposed to explain this pattern, but none has gained general acceptance. For a number of years now, we have been testing existing hypotheses about the origin and maintenance of the LDG and also developing new models. For example, our “Out of the Tropics” model integrates evolutionary processes with biogeographical dynamics to provide new insights into the origin and maintenance of the LDG. [Figure above is from Jablonski et al. 2013]
The latitudinal diversity gradient (LDG) – a dramatic increase in the richness of species and higher taxa from the poles to the tropics – is a global biodiversity pattern shared by both marine and terrestrial ecosystems. Understanding the processes responsible for the origin and maintenance of the LDG has been a focus of biology for over two centuries and is essential for formulating effective management and conservation strategies. Yet, despite considerable progress, the environmental and biological drivers of the LDG still remain poorly understood; two dozen major hypotheses have been proposed to explain this pattern, but none has gained general acceptance. For a number of years now, we have been testing existing hypotheses about the origin and maintenance of the LDG and also developing new models. For example, our “Out of the Tropics” model integrates evolutionary processes with biogeographical dynamics to provide new insights into the origin and maintenance of the LDG. [Figure above is from Jablonski et al. 2013]
Molluscan microbiomes
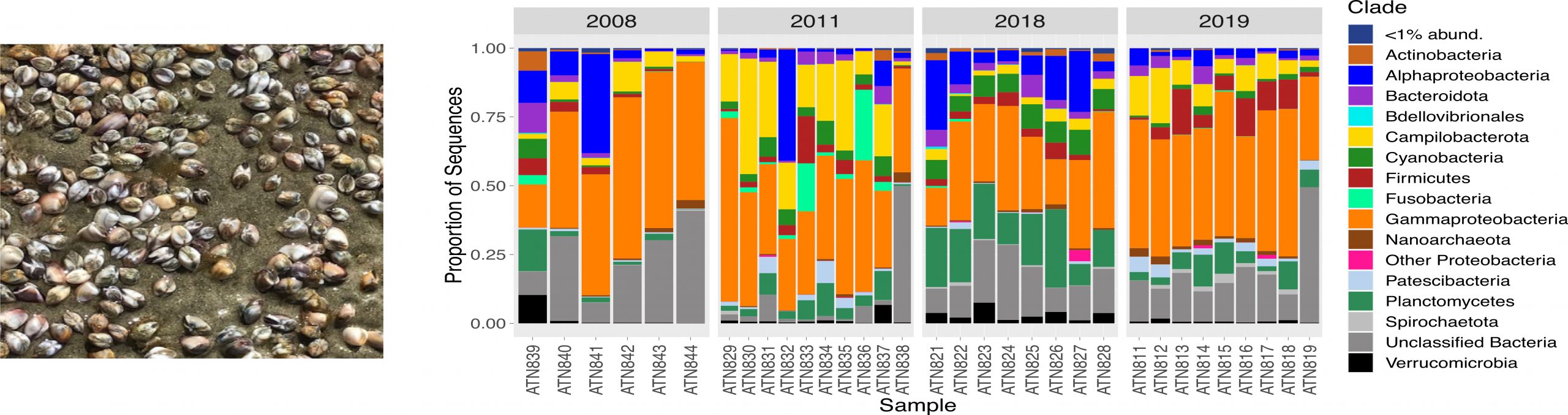
Microbes represent the most diverse and abundant group of organisms on this Planet. They are also known to impact everything from organismal physiologies and metabolism to the functioning of ecological systems and biogeochemical processes. Yet we still know very little about the microbial communities associated with marine invertebrate species. We have recently initiated research projects that will quantify (i) how the microbiomes of marine mollusks vary across large spatial and environmental gradients and the functional consequences of such variation (ii) how such patterns have evolved and (iii) how they are affected by anthropogenic impacts such as coastal eutrophication and climate change. [Figure above is from Neu et al. 2021]
Biotic effects of climate change
 Climate is widely recognized as one of the major determinants of spatial distributions of species but predicting how species and ecosystems are likely to respond to anthropogenic warming remains a major challenge for biology. Our work on the biotic effects of climate change focuses on both ecological (e.g. species range shifts and changes in community compositions) and evolutionary effects. The glacial-interglacial cycles during the late Pleistocene provide a set of natural experiments that can be used to investigate biotic responses to large scale changes in climate. Using a combination of the Pleistocene and Holocene record of living species, molecular markers, geochemical data and phenotypic measurements we are investigating how marine molluscan species responded to past changes in climate. The ultimate goal here is to use this information, in combination with ecological and functional data, to identify species that are likely to be affected by future warming (or conversely those that are likely to be resilient). [For explanation of the figure above see Hellberg, Balch & Roy 2001]
Climate is widely recognized as one of the major determinants of spatial distributions of species but predicting how species and ecosystems are likely to respond to anthropogenic warming remains a major challenge for biology. Our work on the biotic effects of climate change focuses on both ecological (e.g. species range shifts and changes in community compositions) and evolutionary effects. The glacial-interglacial cycles during the late Pleistocene provide a set of natural experiments that can be used to investigate biotic responses to large scale changes in climate. Using a combination of the Pleistocene and Holocene record of living species, molecular markers, geochemical data and phenotypic measurements we are investigating how marine molluscan species responded to past changes in climate. The ultimate goal here is to use this information, in combination with ecological and functional data, to identify species that are likely to be affected by future warming (or conversely those that are likely to be resilient). [For explanation of the figure above see Hellberg, Balch & Roy 2001]
Dynamics of extinctions – past and future
 Human activities are increasingly threatening the future survival of many species, both on land and in the oceans. Emerging evidence suggests that the nature and selectivity of losses due to anthropogenic extinctions may be different from those due to natural causes in the geological past. But we still know little about how human-caused extinctions erode phenotypic diversity and evolutionary histories of clades and how such losses compare to those in the past. We are exploring this issue using simulation models as well as empirical data for marine and terrestrial mollusks. [More on the figure above in Huang & Roy 2013]
Human activities are increasingly threatening the future survival of many species, both on land and in the oceans. Emerging evidence suggests that the nature and selectivity of losses due to anthropogenic extinctions may be different from those due to natural causes in the geological past. But we still know little about how human-caused extinctions erode phenotypic diversity and evolutionary histories of clades and how such losses compare to those in the past. We are exploring this issue using simulation models as well as empirical data for marine and terrestrial mollusks. [More on the figure above in Huang & Roy 2013]
Conservation of coastal ecosystems
Harvesting of coastal marine invertebrates for food and other use is widespread all over the world. Such harvesting is almost always size-selective, preferentially targeting large individuals. However, little quantitative data exist about the biological consequences of such exploitation, largely due to the lack of baseline information against which to compare present day patterns. In most cases we do not know how harvesting impacts life histories and phenotypic traits, whether it leads to extensive local extinctions of the targeted species or changes local abundances and/or populations structures of species. From an evolutionary perspective, preferential loss of the largest individuals in a population represents a novel selective force, something species are unlikely to have experienced in their evolutionary past. So how such harvesting practices will shape future evolutionary trajectories of many species remains an important question. In addition, we also know little about how harvesting affects macroecological relationships or biogeographic patterns. We are addressing these and related issues using coastal marine molluscan species as a model system.